Phototransduktion
Kontakt
Phototransduktion
Forschung in der AG Biochemie Signaltransduktion / Neurosensorische Prozesse
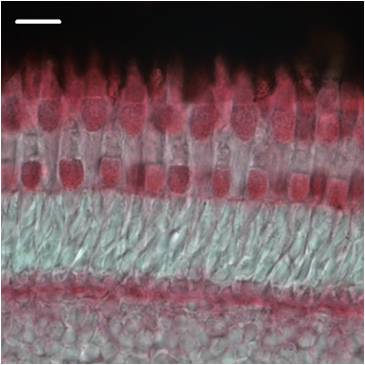
Photorezeptorzellen der Wirbeltiernetzhaut sind exquisite Lichtdetektoren. Mit der Registrierung einzelner Photonen erreichen sie die Grenze physikalischer Messbarkeit. Absorption von Licht und die ersten Schritte der Informationsverarbeitung in der Netzhaut erfolgen in den Außensegmenten der Photorezeptorzellen. Licht wird von dem Sehfarbstoff Rhodopsin (Stäbchen) absorbiert, dieser Vorgang triggert die Aktivierung einer nachgeschalteten Signalkaskade, die innerhalb von einigen hundert Millisekunden den Abbau des intrazellulären Botenstoffes cyclo GMP steuert (Abb. 1). Zeitlich etwas später verringert sich auch die zytoplasmatische Ca2+-Konzentration in der Zelle. Die Konzentration der beiden Botenstoffe cyclo GMP und Ca2+ sind durch mehrere Regelkreise fein aufeinander abgestimmt (Ca2+-Feedback, Abb. 2). Diese für die Zelle wichtigen Regulationsmechanismen werden durch neuronale Calcium-Bindungsproteine wie Recoverin und den GCAPs (guanylate cyclase-activating protein) oder durch ubiquitäre Calcium-Bindungsproteine wie Calmodulin oder S100 b gesteuert und moduliert. So wird die Lichtempfindlichkeit der Photorezeptorzellen (Adaptation) wesentlich über eine Änderung der intrazellulären Ca2+-Konzentration reguliert. Calcium-Bindungsproteine detektieren diese Änderung der zytoplasmatischen Ca2+-Konzentration und übertragen das Signal auf nachgeschaltete Zielproteine. Diese Vorgänge finden an der Oberfläche der in den Sehzellen vorhandenen Disk -oder Plasmamembran statt.
Die Arbeitsgruppe Biochemie versucht, die molekularen Mechanismen dieser Signalweiterleitung zu entschlüsseln und die physiologische Rolle von Calcium-Bindungsproteinen bei der Lichtadaptation aufzuklären. Im Vordergrund stehen dabei Untersuchungen über Protein-Protein-Wechselwirkungen und Struktur- /Funktionsbeziehungen retina-spezifischer Proteinkomplexe. Laufende Forschungsprogramme und geplante Forschungsziele sind in diesem Zusammenhang:
- Interaktionsstudien neuronaler Calcium-Sensoren (Recoverin, GCAPs) und ihrer Zielproteine (Rhodopsinkinase, membranständige Guanylatcyclasen) mit Hilfe von Oberflächen-Plasmon-Resonanz Spektroskopie, Fluoreszenzspektroskopie und isothermer Titrationskalorimetrie
- Strukturaufklärung von Recoverinmutanten und Proteinkomplexen
- Synthese und Durchmusterung von Peptidbibliotheken, um Interaktionsdomänen zu identifizieren
- Isolierung und proteinchemische Charakterisierung bisher unbekannter Photorezeptorproteine; zelluläre Lokalisation und Aufklärung ihrer Funktion bei der Lichtwahrnehmung
Lichtaktivierte Signalkaskade in Photorezeptorzellen
Abbildung 1: Die Signalkaskade der Phototransduktion. Licht löst die Aktivierung von Rhodopsin (Rh zu Rh*) aus, was zum GDP/GTP-Austausch am G-Protein Transducin (T) führt. Die aktive, GTP-gebundene Form von T aktiviert seinen Effektor PDE, der vom inaktiven (PDEi) in den aktiven Zustand (PDEa) wechselt. Aktivierte PDE hydrolysieren cGMP mit hohen Turnover Raten. Die grün markierten Prozesse tragen zur Verstärkung der Kaskade bei. Die Resynthese von cGMP durch die Guanylatzyklase (GC) steht unter der Kontrolle einer Ca2+-Rückkopplungsschleife, an der Ca2+-Sensorproteine (Guanylatcyclase-aktivierende Proteine, GCAP) beteiligt sind. Der Dunkelstrom durch den CNG-Kanal wird von Na+ und Ca2+ getragen, letzteres wird durch den Na+/Ca2+, K+-Austauscher extrudiert, was zu einer Abnahme des zytoplasmatischen Ca2+ führt. Ca2+-gebundenes Recoverin (Rec) hemmt GRK1 (Rhodopsin-Kinase) und verhindert dadurch die Phosphorylierung von Rh*. Details der Deaktivierungs- und Regenerationsprozesse von Rh* sind nicht dargestellt (modifiziert nach Koch und Dell'Orco, 2015).
Ca2+ -Feedback
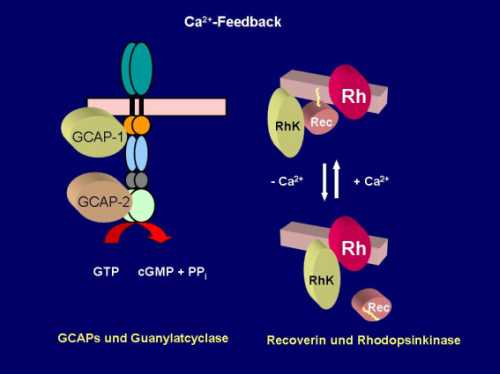
Abbildung 2: Das Absinken der intrazellulären Ca2+-Konzentration nach Belichtung wird von Ca2+-Sensorproteinen detektiert. Dazu zählen insbesondere GCAP-1 und GCAP-2, die zwei in Sehzellen vorkommene membranständige Guanylatcyclasen bei niedrigen Ca2+-Konzentrationen aktivieren. Ein anderer Ca2+-Sensor ist Recoverin (Rec), das bei hohen Ca2+-Konzentrationen die Rhodopsinkinase (RhK) inhibiert. Sinkt die Ca2+-Konzentration, dissoziiert der inhibierte Komplex, Rhodopsin (Rh) wird phosphoryliert und dadurch abgeschaltet.
LITERATUR:
Koch, K.-W. and Dell'Orco, D. (2013) A calcium relay mechanism in vertebrate phototransduction. ACS Chem Neurosci. 4, 909-917.
Koch KW (2013) The guanylate cyclase signaling system in zebrafish photoreceptors. FEBS Lett. 587, 2055-2059.
Koch, K.-W. and Dell'Orco, D. (2015) Protein and signaling networks in vertebrate photoreceptor cells. Front. Mol. Neurosci. Vol.8, Article 67